Some Preliminary Considerations toward
Development of a Mathematical Model of the
Autogenic Brain Discharge as Spontaneous
Localization in Quantum Measurement©
(circa September of 1980)
WILLIAM L. PENSINGER
DOUGLAS A. PAINE
(Suggestion for McIntosh users: Using Virtual PC
with IBM-platform browsers will insure
text-embedded math symbols are read properly.)
TABLE OF CONTENTS Abstract Introduction 1. The Question of the “Classical Limit” 2. Quantum Theory Under Non-linear Reinterpretation for Biological Systems 3. Quantum Determinism and HETEROstasis 4. Evidence Supporting Complementarity in Biological Processes 5. Psychosomatic Interaction and “Relative State” Activation of the Brain 6. A Wave STATEMENT for an Autopoietic Process 7. DNA-Mediated Quantal Localization and Fusion in Cortical Function and Hyden's Molecular Theory of Memory 8. Quantum Measurement, Autogenic Discharge, and Biological Information Exchange at Critical States References Figures Appendix Wherein a Mathematical Model of a Psychosomatic System is Presented in the Cross-notation of G. Spencer Brown's Calculus of Indications
Introduction
Any attempt to formulate a mathematical description of the autogenic discharge must be based upon a hypothesis as to the fundamental role of autogenic discharge activity in organization of a biological process. The most likely hypothesis would be that drawn clearly in relation to the accumulated clinical findings and experimental data associated with neuropsychological application of autogenic therapy. Consideration of the following questions will be helpful in this regard, particularly as many of the principles of the autogenic model of disease (which have evolved by revision through clinical application since the 1920s) are at odds with those of the prevailing mainstream remedial medical model. By what process does neuronal material (e.g., neuronal records of traumatic experiences and unfulfilled needs) acquire a pathofunctional potency [1, pp. 118-123 and 130-133] which sufficiency disturbs homeostatic mechanisms so as to require neutralization [2, 3] through autogenic brain discharge activity? An essential feature of the autogenic model of disease is etiological facilitation by pathofunctional potency of neuronal material. This etiological facilitation, according to the autogenic model, is a contributory factor in onset, and subsequently in course, of all diseases. How are pathogenic effects of this neuronal material communicated to various organizational levels of the living system, and, conversely, how are therapeutic effects of autogenic discharges [1, 130-133 and 2, pp. 1-5] communicated throughout the same hierarchy? Autogenic therapy relies most essentially on practice of the basic autogenic training exercises, which involves internal verbalization of training formulas. Through what mechanism(s) does the manipulation of attention (i.e., the psychological process of passive concentration [1, pp. 11-17] to prescribed psychophysiologic structures) promote the conditions required for initiation of autogenic brain discharge activity, with its demonstrable physiologic effects? Once the autogenic neutralization process has been initiated, by what mechanisms do neutralization antagonizing forms of resistance (NAFR) [3, pp. 4-32] disrupt the therapeutic effects of autogenic discharge activity?
These unanswered questions elaborately illustrate the fact that any mathematical model of the autogenic discharge must: [1] directly represent the interaction between psychological and physiological processes (based upon presentation of some explicit formalism regarding this relationship); [2] take into account the hierarchical nature of the organism and the well-documented multiscaled effects of autogenic discharge activity; [3] provide a representation of the autocatalytic nature of the processes involved. But where is the autogenic theoretician to turn in seeking the required ideas upon which to base such modeling efforts? Contemporary theories of brain function have proved themselves inadequate. Contemporary theories of immune system function have proved themselves inadequate. Contemporary general systems theories of hierarchical organization have proved themselves inadequate. Contemporary notions of autocatalysis, drawn largely from stereochemical metabolic-pathway models, have proved themselves inadequate. This inadequacy is most lavishly apparent relative to degenerative disease. When practice of the standard exercises of autogenic training is demonstrated to be associated with remission effects on course of systemic lupus erythematosus (personal communication from W. Luthe, 1974, regarding clinical trials conducted by T. Abe at Tokyo University), it is abundantly clear that the autogenic theoretician cannot rely exclusively upon the biology and chemistry underlying the remedial medical model, which provides so little insight into the nature of degenerative disease. The present paper will present a perspective on the questions raised above, which involves application of a non-linear reinterpretation of quantum principles to theory of biological organization, based upon the hypothesis that the autogenic brain discharge fulfills its fundamental biological role by being equivalent to spontaneous localization in quantum measurement. This notion regarding brain discharge and quantum measurement may, when first encountered, appear incongruous. Consideration of the likelihood that sub-clinical autogenic discharges are an integral part of adaptation (incorporating normative growth and repair processes) to a changing environment and that all perception and proprioception involves environmental scanning (i.e., “measurement”) may suggest that, in fact, there is no such incongruity. Moreover, from perspective of a nonlinear treatment of the quantum wave function, that wave function�s collapse in spontaneous localization exhibits features of discharge phenomena.
Immediately, the autogenic theoretician is confronted with the question of functional integration. There is elaborate documentation clearly demonstrating that autogenic discharge activity is associated with a prohomeostatic shift in the functioning of a wide variety of psychophysiologic processes [4, 5]. The autogenic discharge most certainly interacts in some manner with those mechanisms maintaining functional integration. But contemporary stereochemical models of functional integration have proved themselves inadequate. The mechanisms governing functional integration in living systems are not well understood. Were this not the case, degenerative disease would not be the mystery it is. Were degenerative disease not mysterious, its treatment would not be so unsuccessful as it is under the remedial medical model. Such a circumstance suggests that study of stereochemical relations is not sufficient to provide the clues necessary for adequate understanding of autogenic discharge phenomena, and that the autogenic theoretician is well advised to look elsewhere -- to re-interpretations of quantum theory, for instance. Spontaneous localization, under non-linear treatment of the wave function, is not to be conceived as the sudden appearance of mere discreteness, mere simple-location, and mere absolute separability -- as has been the case in the standard linear treatment of the wave function and its collapse. Non-linear spontaneous localization at collapse of a time-differentiated wave function is a radiational discharge conveying nonlocal, functionally integrative, holographic-type information into arrays of functional specificity, which (arrays) may be regarded as structures in exactly the manner that automatized functions are regarded as structures in Heinz Hartmann�s conception of automatization and deautomatization presented in his well known classical account of the problem of adaptation.
Functional integration is most essentially a question of scale interaction. How are the multitudinous relations between submolecular quantum processes, molecules, macromolecules, cells, tissues, and organs coordinated into the unified purposeful activity which constitutes an organic whole? No one knows, as contemporary theories of scale interaction have proved themselves inadequate, deeply implicating the thermodynamic (heat-driven) stereochemical model of multiscale metabolic processes. Multiscaled processes are poorly understood not only in the biological sciences, but also in physics where scale interaction, cooperative and critical phenomena, and
n-body problems remain frontier areas of research.
There also are no clear guidelines regarding psychosomatic interaction, as contemporary approaches to the Leib-Seele problem have proved themselves inadequate. The question of the relationship between the higher psychological functions and the physical substrate remains so much an area of controversy in the neurosciences one suspects the mind substrate to the physical body as epiphenomenal artifact of enculturation. In the 1950s, the prevailing point of view held that the brainstem reticular and centrencephalic systems are the fundamental physical seat of consciousness [6-8]. There has recently been a shift of opinion, partially the result of split-brain research, where the physical organ mediating awareness is thought to be the neocortex [9-15]. Wilder Penfield, however, in 1975 [16], argued that the centrencephalic system must be of primary importance. And very significantly, the controversy has spilled over into the area of quantum theory, where the measure problem (the question of observer-state object-system interaction) is often seen as analogous to the Leib-Seele problem [17-26]. One wants to know if the autogenic discharge is the result of an observer-state object-system interaction (which, in the standard interpretation of quantum mechanics, leads to a collapse in the wave function, with associated spontaneous localization) in the form of self-observation during practice of the autogenic standard exercises [27, pp. 23-141]. Experiments could be designed quite specifically in relation to this issue, given that it has recently been demonstrated by research conducted in Budapest (description of research, Autogenic Therapy Symposium, 4th Congress of the International College of Psychosomatic Medicine, Kyoto, August, 1977) that differences in the anti-epileptic effects of the autogenic training exercises practiced on various discharge levels (as viewed on discharge histogram) relate to different sectors of the circadian cycle. (It is noted that this piece of experimental data is particularly relevant to the idea of an “intracellular Zeitgeber” which emerges from the Paine-Pensinger model of superconductant DNA, particularly such a Zeitgeber in neural and perineural tissue.) But to which of the many interpretations of quantum mechanics is the autogenic theoretician to subscribe? Most of the interpretations [28-34], though they may begin in novel fashion, become experimentally indistinguishable from the von Neumann interpretation [35]. This interpretation carries with it an implied concept of mind-body interaction. To quote von Neumann:
… it is a fundamental requirement of the scientific viewpoint - theBut the autogenic theoretician cannot at all take this perspective seriously, as it completely rules out application of the von Neumann interpretation of quantum mechanics to the quantum level of functioning of human brain. The von Neumann interpretation relies on Hilbert space (a hyperdimensional function-space) to represent quantum interactions of physical particles. The requirement that processes of subjective perception must be referenced only to “ordinary space” of three ponderable dimensions means that, by the von Neumann model, physical processes alone can take place in non-ordinary hyperspace, not also psychological processes. The world of immediate experience is thus turned upside down by von Neumann: he would have us believe that the hard physical objects seen in everyday life are rooted in non-ordinary hyperspace, while processes of the mind are rooted only in ordinary space wherein no mental objects are visually apprehended. Is it possible that the current hiatus in quantum theory is intimately related to the current paradigm crisis [36] in psychiatry and medicine?
so-called principle of the psycho-physical parallelism - that it must be possible so to describe the extra-physical process of the subjective perception as if it were in reality in the physical world - i.e., to assign to its parts equivalent physical processes in the objective environment, in ordinary space. [Emphasis added, 35, pp. 418-419.]
The Question of the “Classical Limit”
Consider the proposition that major progress in these areas is blocked by one fundamental assumption: that there is a “classical limit” beyond which quantum mechanical principles do not apply. It is everywhere assumed that the classical Newtonian laws of physics govern events in the middle scale levels of reality - our world of everyday experience. In awareness of this conventional assumption, let us examine the “Relative State Formulation of Quantum Mechanics” [37] as presented in a paper published in 1957, which was written by Hugh Everett, III. The theory developed in this article and a longer one entitled “The Theory of the Universal Wave Function” [38] has become known as the “many-worlds interpretation of quantum mechanics” [32, 39]. We will discuss this formulation at some length, because, in modified form, it will play a central role in the ideas to be developed later in the present paper. Everett�s perspective advocates a direct correspondence between the mathematical formalism of quantum theory and the structure of reality [19, 40]. It assumes that the state function never collapses, and that, as a result of the interactions between an object-system and an observing-apparatus, the universe is continually branching into a multitude of mutually unobservable and independent domains, each of which is no less real than any other. The theory is completely deterministic, and does not, therefore, rely on probability concepts, as does the standard interpretation. Because the branches of the state function do not interact (and, hence, no independent domain can observe any other independent domain) the observer cannot be aware of the splitting process [19]. In spite of the unusual world construct the theory gives rise to, the analysis of the measurement process (which relies heavily on the von Neumann formalism) yields an interpretation experimentally equivalent to the conventional probabilistic conception.
When Everett wrote these papers his purpose was not to establish a new departure in quantum theory, but, rather, to provide a general theoretical framework from which the standard interpretation could be derived. There was no doubting on the part of Everett that our usual experience of reality might in some fashion be misleading. This lack of doubt was formulated into THE critical transition in the argument when shifting to the von Neumann formalism wherein it was shown that the multi-worlds interpretation is experimentally equivalent to the standard probabilistic conception. In considering the behavior of macroscopic objects resulting from the superposition of quantum states, Everett says:
Immediately after this passage, wherein an analogy to Schrödinger�s cat is given relative to a cannonball, Everett introduces the von Neumann formalism and proceeds with the elaborate complexities of argument which eventually demonstrate experimental equivalence of the many-worlds interpretation and the conventional probabilistic conception. He provides an analysis of the measurement process, where the branches of the state function do not interact, in order to show how this difficulty with the cannonball's behavior can be overcome, thus yielding the standard interpretation.Suppose, for example, that we coupled a spin measuring device to a cannonball, so that if the spin is up the cannonball will be shifted one foot to the left, while if the spin is down it will be shifted an equal distance to the right. If we now perform a measurement upon a particle whose spin is a superposition of up and down, then the resulting total state will also be a superposition of two states, one in which the cannonball is to the left, and one in which it is to the right. There is no definite position for our macroscopic cannonball!
This behavior seems to be quite at variance with our observations, since macroscopic objects always appear to us to have definite positions. (Emphasis added.) Can we reconcile this prediction of the purely wave mechanical theory with experience, or must we abandon it as untenable? [38, pp. 61-62.]
But can we be so sure that “macroscopic objects always appear to us to have definite positions”? Is it possible that our unquestioned assumption that objects always have definite positions is the result of an enculturated avoidance of dissociative experience, this avoidance being an issue of central concern to the prevailing crisis in psychiatry? An alternative interpretation would advocate the notion that the “definite position” of any given object (macroscopic or microscopic) at any given scale level of reality contains a superposition of information, which, upon being decoded, would provide a holographic map of the object�s interactivity with its nested and nesting environments on other scale levels of reality. This holographic map would exist as a frequency domain, or superpositioning of “quantum-measurement” time frames, defining properties of the object in question - in a manner analogous to how the superimpositions of the Analytical Cubist image characterize the object of a Cubist painter�s perception. Interactivity of “independent” domains would be mediated by the stacked time frames the domains are subject to. This notion will be discussed at some length later in the present paper.
Recently, Bernard D�Espagnet [33] and B. J. Hiley [41] have raised the question of relevance to quantum theory of Piaget�s finding [42, 43] that the concept of simply-located separable objects is a result of the process of cognitive development. The finding that �object constancy� arises at a particular ontogenetic stage of development of human cognitive function casts doubt on the proposition that objects are only simply-located, and, hence, that there must be only one state of the state function that is actual relative to a macroscopic object.
Of even greater importance in this regard is work presented us seven years before Everett�s paper appeared. In 1950, posthumously, the JOURNAL OF THE OPTICAL SOCIETY OF AMERICA [44] published the results of studies made by Rudolf K. Luneburg at Columbia University�s Knapp Memorial Laboratory of Physiological Optics into the “… sensed qualities of form and localization in the three-dimensional continuum of visual perception” [44, p. 627]. These studies experimentally demonstrated that visual space is a non-Euclidean metrical space, the distance function of which varies with psychological factors and depends upon the individual observer. Hence, Luneburg coined the term “psychometric distance function”. Three years later, published in the same journal, a paper [45] written by one of Luneburg�s associates, Albert Blank, maintained that when the effects of motion are considered, a spacetime metric is required, which has a visual limiting velocity and Lorentz-Fitzgerald contraction associated with it. Luneburg�s experiments demonstrated that physical objects can be localized in the visual field only through use of cognitively learned cues. In his words:
Visual localization of an isolated, repeatedly shown physical point P* is found to be highly erratic. The point P seems to be floating in space indeterminedly, now it seems near, now far. [44, p. 628.]Later in the paper, he summarizes:
These observations are concerned with the thesis that there is no absolute localization even in binocular vision. (Emphasis added.) A given physical configuration provides no immediate visual clues which could determine localization and absolute phenomenal size of the sensed configuration. Localization and size are determined by other non-visual psychological factors, in particular by the intellectual experience of the observer. [44, p. 631.]
This evidence suggests a fundamental reorientation of thought in many areas, but particularly so with regard to the question of collapse of the wave function in quantum measurement. Everett says: “… macroscopic objects always appear to us to have definite positions.” Luneburg says: “… there is no absolute localization even in binocular vision.” Who are we to believe? Given that Luneburg�s experimental findings involve visual space having the non-Euclidean spacetime metric and Lorentz-Fitzgerald contraction of the Special Theory of Relativity, it appears that relativistic factors are directly involved in the quantum measurement process. Thus, in the present attempt to find a framework within which to understand autogenic discharge activity, we will not assume the standard interpretation of quantum mechanics or existence of a “classical limit”. On the contrary, we will assume that quantum principles apply to all scale levels of reality.
Quantum Theory Under Non-Linear Reinterpretation for Biological Systems
In the following, a general conception of the relevance of quantum principles to macroscopic biological processes will be presented which establishes a new perspective on functional integration and biological information exchange. This perspective provides a context within which to explore the relevance of cooperative and critical phenomena to biological information systems and suggests a new conception of psychosomatic interaction. In this context, a hypothesis may be formulated concerning the fundamental role of autogenic discharge activity in organization of a biological process.
Let us consider the supersystem-system-subsystem structure [46-48], which constitutes the hierarchical organizational framework of a macroscopic biological process, to be governed by quantum principles. Thus, we would have a macroquantum system which can be represented by a state function. All of the whole integers values of the state function represent interaction quanta (e.g., temperature perturbations) arising within the superposed supersystem-system-subsystem framework. And the description of the total configuration of such quanta, each represented by a given eigenvalue having real and imaginary components, is a deterministic description of the result of all possible interactions of an element of the hierarchy with its embedding supersystems and embedded subsystems. Likewise, the complex nonlinear interaction, both within and outside the embedded domain, is represented by superposed eigenfunctions having real and imaginary components.
In our view of a macroquantum system, a given fundamental frequency established for an element of the hierarchy represents a state of interaction between this macroquantum domain structure [46] and a given level of its embedding hierarchy. The multiple values of the state function are not interpreted as representing a probabilistic wave function, but, rather, as representing the interactivity derived from a fundamental frequency. This, in turn, deterministically governs functional integration of specific single-valued processes constituting the supersystem-system-subsystem hierarchy - which is to say that the microphysical structure of the organism functions as a multivalued referencing frequency domain [49-53] for the hierarchically organized information exchange process. The single-valued information that is exchanged between macroquantum domain structures on the molecular, macromolecular, and cellular levels (i.e., through stereochemical metabolic, hormonal, hematopoietic, lymphatic, muscular, neuronal, and immunological mechanisms) is referenced to the multivalued frequency domain. The quantum structure of the organism thus serves a functional integrative role, and, so to speak, keeps a running multivalued, composite accounting of all single-valued information exchanges on other scale levels.
The associated Schödinger equation would, therefore, be reinterpreted within such a context. As Everett has argued that it is meaningful to suppose a state function for the whole universe [40], so we believe it is meaningful to suppose state functions for the organism as a whole, and given macroquantum domain structures. In Everett�s interpretation, the constituent states y1, y2, y3, …,yn) of the state function (Y) are all considered to be actual. Likewise, we hypothesize that, in the macroquantum description of a biological information process, the constituent states of the state function are all actual. However, this supposition, in the present biological context, leads to quite different conclusions than Everett has derived concerning fundamental questions in quantum mechanics per se.
The change of state of a given macroquantum domain structure, defined in terms of phase boundaries and considered as an isolated system, transpires in a manner described by the linear wave equation [37]
where A is a linear operator. This change of state is deterministic and continuous, so long as the domain structure remains in an isolated state. However, since there is constant information exchange taking place between the different hierarchical levels of organization within the organism, and between the organism and the environment, the macroquantum domain structure cannot be considered an isolated system. Rather, the domain structure must be undergoing a rapid succession of information exchanges. It is as a result of these information exchanges, and the underlying scale interaction process, that the state function of a macroquantum domain structure collapses. Under reduction of the state function, the change of state transpires discontinuously in correlation with the discrete sequence of interaction quanta received by the domain structure. In order to present a mathematical description of these concepts, we first note that, in the biological context, it is not possible to understand why the state function collapses if the system is considered to be isolated. This collapse cannot be fully understood even in an open system. Collapse of the state function becomes understand-able only in a self-referential system undergoing multiple scale interactions and self-reentrant autocatalysis. While the linear time-dependence of state of an isolated system is governed by the linear wave equation, the linear-time independence of state of an open system undergoing multiple scale interactions is governed by a second order wave equation
where B is a nonlinear operator. The linear-time independence of state of a self-referential system undergoing self-reentrant autocatalysis is governed by a third order wave equation
where C is also a nonlinear operator.
Before going further, let us examine the linear-time dependency and linear-time independency (but nonlinear-time dependency) using terms of an equation derived by the authors to explain the dynamics of superconductant DNA molecules [54]. By letting t = (To - Tc), or the temperature interval between the DNA molecule�s reference level and critical temperatures, the following expression was obtained for evaluating information definition and exchange by DNA:
where P, Q, and R are nonlinear- and linear-time dependent coefficients. In term #1, the quantum of information to be exchanged is defined by the linear-time taken by a parcel of p-electrons in transiting To«Tc. The inverse of this period establishes the fundamental frequency (no) of this quantum. The number of such transits establishes the number of quanta to be read. Term #2 evaluates the linear-time taken to receive these quanta (which themselves exist as a sinusoidal function of linear-time) within the structure of the DNA molecule. The double helix of the molecule spatially encodes this frequency shift once the quantum signature has been incorporated within the DNA molecule�s structure. Thus, the constraining force interpreted by J. L. Synge�s examination of “The Electrodynamic Double Helix” [55] becomes, from our point of view, the scale-interactivity dictated by the nonlinear wave equation presented in (2). Term #3 evaluates the nonlinear-time taken by the embedding environment to receive the signal represented by term #2. Clearly, both of these information exchanges represent higher-order time differentiation with regard to the initiating or linear-time dependent signal. And while the signal itself is irreversible, the ability to erase (nullify or cancel) the signal within the DNA molecule or its environment is reversible.
Quantum Determinism and HETEROstasis
In the present biological context even the reduced modality of the state function is deterministically governed, but the deterministic march is transverse to linear-time. The reduced modality is a decomposed version of the composite modality. In the composite mode, the constituent interactivity states are given simultaneously so that a nonspatial and linear-time independent interactivity spectrum is represented. The whole supersystem-system-subsystem hierarchy is portrayed in a non-hierarchical, superposed, multivalued, instantaneous holographic fashion. In the reduced mode, the multivalued composite is decomposed into a supersystem-system-subsystem hierarchy represented in a spatial and linear-time dependent context, where the constituent interactivity states are presented sequentially as if the domain structure were undergoing information exchanges with different levels of the hierarchy one after another. In other words, in the reduced mode each constituent state of the state function represents a state of interaction between the given domain structure and some other element of the system hierarchy. In the composite mode, the total array of constituent states of the state function associated with the given domain structure represents the interactivity state of the whole organism, portrayed through the interactivity spectrum of the given domain structure. This mirroring of information between the whole and the part is the result of a “relativity of states” between the elements of a system hierarchy. To quote Everett:
Because of the information reciprocity between the composite and decomposed modalities, the collapse of the state function and its recomposition must be considered a self-reentrant autocatalytic process.There does not, in general, exists anything like a single state for one subsystem of a composite system. Subsystems do not possess states that are independent of the states of the remainder of the system, so that the subsystem states are generally correlated with one another. One can arbitrarily choose a state for one subsystem, and be led to the relative state for the remainder. Thus we are faced with a fundamental relativity of states, which is implied by the formalism of composite systems. [37, p. 456.]
It is this oscillating, self-reentrant autocatalytic phase transition, postulated to establish a governing reference frequency necessary for the control of growth and repair processes, which is hypothesized to mediate the requirements of functional integration and specificity in hierarchically organized organic systems. It is not that the operators corresponding to the dynamic variables represent all possible states of the system, one of which is actual, but, rather, that the operators represent the simultaneous interaction of the domain structure with all actual elements of the supersystem-system-subsystem composite - this representation being necessary for the functional integration of the organism. Collapse to the single-valued sequential representation is necessary to meet the needs of functional specificity.
Furthermore, as the scale interactions undergone by the macroquantum domain structure (and the oscillating phase transitions between the composite and reduced modalities of the state function) are thermodynamically reversible (transverse to linear-time), interference patterns develop between the various branches of the state function. Interference between two branches represents an information exchange between two domain structures. The total configuration of these interference patterns constitutes a map of the overall state of scale interaction manifest in the supersystem-system-subsystem hierarchy. It is for this reason that all of the branches of the wave function are retained and regarded as actual.
When the system is viewed from perspective of the multivalued modality, the system composite is not characterized as decomposed into its single-valued hierarchy. Scale interactions from this perspective are mapped as continuous branch interference patterns in the manner of interference patterns on a holographic plate. After the phase transition, wherein the multivalued composite is characterized as decomposed to the modality of the single-valued hierarchy, scale interactions are mapped as discrete quanta of free-energy packets exchanged between macroquantum domain structures on all scale levels of the system hierarchy. The modality of continuous interference patterns underlies the mechanisms fulfilling requirements of functional integration, while the modality of discrete quanta exchange underlies the mechanisms fulfilling requirements of functional specificity. The sum total of the information contained in the multivalued composite is identical to that contained in the decomposed single-valued hierarchy. The same information exists in complementary modes of representation.
There is another form of complementarity build into the macroquantum description of a biological process, as already suggested by the superconductant DNA model: that between reversibility and irreversibility. Because the state function in its composite modality contains information solely in multivalued form, this modality is the linear-time independent mode of the system. Linear-time dependence requires a single-valued sequence of self-identical instants. Thus, the decomposed single-valued modality is the linear-time dependent mode of the system. Transitions between the linear-time independent and dependent modes are reversible, while variations of the linear-time dependent mode are irreversible. The continual transitions between linear-time independent and dependent modes drive variation of the linear-time dependent mode, because the reversible transitions govern free-energy flow through the system hierarchy.
In other words, variations of the linear-time dependent mode are a function of oscillatory parameters governing the repeating transitions between composite and decomposed modalities. As efficiency of this linear-time independent oscillatory process increases, the linear-time dependent variation tends toward a state of maximal free-energy. This maximal state has been termed heterostasis. The concept of heterostasis is an important contribution to biological thought formulated by A. H. Klopf. To quote from Brain Function and Adaptive Systems: A Heterostatic Theory:
Nervous systems are so structured that homeostasis is a necessary (but not sufficient) condition for the maintenance of heterostasis… An organism will be said to be in a condition of heterostasis with respect to a specific internal variable when that variable has been maximized. Heterostasis, as the term itself suggests, is not associated with a steady-state condition. In general, the internal variable to be maximized will have an upper limit that changes as environmental constraints change. Therefore, even if heterostasis is continuously maintained by an organism, a steady-state condition with respect to the internal variable will not result. Furthermore, maintenance of the condition of heterostasis is not necessary for survival, in contrast to maintenance of homeostasis which is an essential condition for life. With respect to heterostasis, it will in fact be seen that living systems infrequently or never achieve the condition. For this reason, it is appropriate to define a heterostatic system as one that seeks (moves toward) a maximal condition while not necessarily ever achieving it. [56, pp. 5 and 8.]Heterostasis is that state of a biological system toward which irreversible processes move as the efficiency of the governing reversible process increases. This is to say, as efficiency of scale interactions in the supersystem-system-subsystem hierarchy increases, the governing oscillation drives the system through a sequence of phase transitions toward heterostasis.
Evidence Supporting Complementarity in Biological Processes
If these quantum principles do apply to macroscopic biological processes, then there must be considerable experimental evidence and theoretical perspectives in the existing scientific literature to suggest that this is, indeed, the case. In an earlier paper [57] the present authors proposed that sufficient evidence exists to hypothesize that two basic states of organization are inherent in hierarchically organized organic systems. The current context of a macroquantum description suggests that these two basic states are analogous to those indicated by the principle of complementarity in quantum theory. The localized “particle”-like aspect of a biological system would be those structures and processes governed by the principles of stereochemistry. But what about the nonlocal “wave”-like aspect of a biological system? Is there any evidence to suggest that macroscopic organic processes manifest this complementary “wave”-like modality?
There appears to be considerable evidence. Research in the area of biological effects of electromagnetic fields has proceeded along many fronts for nearly fifty years. In the early 1930s, Yale anatomist H. S. Burr began to measure voltage gradients in various plant and animal organisms and to correlate these gradients with anatomical and physiological properties [58-63]. In 1935, he published, along with F. S. C. Northrop, an important paper describing an electromagnetic theory of life [64]. His later papers include studies of the electromagnetic aspects of malignancy and wound healing
[65-67]. In the past three decades there has been an accelerating interest in all aspects of this area. The electrical properties of tissues have been studied [68-72]; research on photoelectric [73], pyroelectric [74, 75], and piezoelectric [74, 76] properties of organisms has been conducted; in vivo studies have demonstrated that alterations in the electromagnetic environment are capable of influencing growth and repair processes (e.g., osteogenesis, limb regeneration, wound healing, carcinosarcoma regression, and cartilage repair) [77-91]; studies of the effects of electromagnetic forces on living organisms have led to the hypothesis that there is a phylogenetically earlier analog data system to complement the developmentally later digital system [82]; studies of magnetic fields surrounding the human head (MEGs) and torso have been made [92-95]; it has been determined that active nerves emit electromagnetic fields [96, 97]; it has been suggested that electrical activity at synaptic junctions form neural holograms [98]; evidence has accumulated to suggest that the periodicity of endogenous biological clocks is affected by fluctuations in the environmental electromagnetic field [99-104].
Paralleling these investigations, there has been increasing research within the field of quantum biochemistry into the role of charge transfer interactions which implies that quantum nonlocality may be a principle of fundamental importance in the organization of living systems [105, 106]. Though, as M. A. Slifkin says, “There is no clear evidence of any role played by charge transfer complexes in biology” [ 106, p. XIX], little doubt can be entertained that charge transfer processes will be found to exercise control functions in metabolic processes. R. S. Mulliken was one of the first to use the term “charge transfer complex”[107, 108] and A. Szent-Györgyi, over a period of twenty years, has written a series of books describing the likely role of charge transfer interactions [109-112] in mediating normative (necessary to growth and repair processes) shifts between “alpha” and “beta” states, conceived as phylogenetically earlier and later in evolutionary origin. He has also proposed a theory of cancer based upon the principles elaborated in these books [113], wherein carcinogenesis is conceived as resulting from the organism�s loss of ability to make repeating transitions between the “alpha” and “beta” states -- the involved transitional oscillation being what the present authors describe as a reversible “linear-time independent oscillatory process” governing linear-time dependent stereochemical variations.
In the field of psychology there are also many indications of a complementarity of functional modes paralleling that exhibited by the physiological aspects of the organism. Freud was one of the first to draw attention to this dichotomy when he distinguished between “primary process” and “secondary process” functioning [114, 115]. Later developments of Freud�s formulations - particularly in the structural theory of ego psychology [116], the theory of attention cathexis [117], and the metapsychology of hypnotic induction and the hypnotic state [118] - gave rise the theory of automatization and deautomatization [57, 115-119], which requires two distinct functional modes. When, through self-observation, attention cathexes are directed toward automatized sensory functions, a deautomatization occurs, which is frequently accompanied by a fusion of the modalities of sense perception. Synesthesia [120] and eidetic imagery [121] both support psychological complementarity.
The work of Leonard J. Ravitz, an associate of H. S. Burr and F. S. C. Northrop, is of particular significance in the present context. In 1950, Ravitz published in Science [122] an account of his electromagnetic field monitoring of the hypnotic state. Later publications [123-127] elaborated upon his findings, which he summarizes in the following statement:
Briefly, electric and clinical evidence suggest that hypnosis represents a change of state function involving shifts in the balance of the phylogenetically ancient basal ganglia with respect to the phylogenetically recent neocortex, in which field properties of the entire organism participate. [127, p. 17.]This formulation concerning an altered state of consciousness is strikingly similar to the prevailing theoretical perspective in autogenic therapy concerning the autogenic state and autogenic shift, where it is “… postulated that the autogenic state promotes a regressive tendency of the activity of phylo- and ontogenetically younger parts of the brain, and a corresponding activation of developmentally older parts of the central nervous system” [128, p. 31].
This considerable accumulation of evidence clearly suggests that biological systems, both in physical and psychological aspects, conform to the quantum mechanical principle of complementarity. The one modality is primarily concerned with functional specificity; the other, with functional integration. In Figure 1, a schematic representation is provided of the concepts involved. It is hypothesized, as discussed above, that the system oscillates (in a linear-time independent fashion) between the composite and decomposed modalities. The transition from the single-valued to the multivalued mode is termed a “critical phase transition” because, in order for this recurring process to mediate scale interaction in the system hierarchy, there must be a transit from a reference state to a critical state, and return.
In the theory of critical behavior [129, 130], it is found that a physical system near its critical point develops fluctuations at all possible scale levels. Normally, elements of a system remain uncorrelated that are associated with scales of organization greatly removed from one another in size. But as the system approaches its critical state, it is found that the coupling strength of critical variables increases as correlations across a wide range of length scales are established. At the critical point, the correlation length is found to be infinite and critical variables are coherently coupled across all scale levels of the system. Examples of critical multiscale phenomena include macromolecular conformation, spontaneous magnetization, and superconductivity.
Psychosomatic Interaction and “Relative State” Activation of the Brain
Oscillation between composite and decomposed modalities is reflected in both psychological and physiological aspects of the organism. The “functional disintegration“ or “splitting up for release” of neuronal material during autogenic abreaction [2, p. 4] is an expression of transition from composite to decomposed modalities: a multivalued structure is split into its separate single-valued functions (in exactly the manner described in Hartmann�s account of adaptation through deautomatization and reautomatization, wherein a structure is treated as a function automatized) [116]. The fusion of sense modalities leading to synesthetic experience (e.g., colored hearing) is an expression of the inverse process: separate single-valued contents are fused into a unitary multivalued content. These reciprocal processes are the psychological aspect of an underlying physical process whereby information in the system hierarchy continuously undergoes a macroquantum process of spontaneous localization and fusion [46]. It is the localization and fusion process that mediates psychosomatic interaction in living systems.
But what does it mean to say that there is a process of spontaneous localization and fusion occurring in a biological system? The brain may receive inputs from as many as 106 sensory receptors simultaneously, while digital computers handle only one at a time [131]. Because of this fact, it has often been suggested that the brain may be equivalent to many computers connected in parallel so as to be able to deal with the large number of simultaneous inputs. But is it not more reasonable to suppose that the organism engages in a processing of multivalued information, rather than parallel processing of single-valued information? The quantum structure of the organism, by virtue of superposition, is a multivalued processor.
Current concepts concerning the relationship of mental functions to physical processes are based largely upon research that has been conducted on the brain stem reticular system [132], the architecture of the cerebral and cerebellar cortex [139-144], studies of functional localization and of the higher integrative functions of the brain (i.e., the hypothesized centrencephalic system) [7, 8, 16, 145-149], and split-brain research [13-15, 150-156]. While it is far beyond the scope of this paper or the competence of its authors to summarize this vast area of research, it is clear that these investigations have generated approaches to the problem of what makes the unitary stream of consciousness possible, all of which assume that the physical substrate of awareness is to be localized in the brain. Penfield and Jasper [7] attribute the overall integration of the elements of sensation into sensory images and patterns of abstraction and memory to a centrencephalic system localized in the higher brainstem. Eccles [10, 11] has described the mind-body problem as having reduced itself to a question of the relationship between mind and the linguistic liaison areas of the dominant cerebral hemisphere, where the patterns of module interaction, design, and operation are of crucial importance. Sperry [13, 15, 156] as suggested that consideration of the findings associated with surgical section of the cerebral hemispheres leads to the conclusion that the higher psychological functions cannot be associated with the brainstem reticular and centrencephalic systems. Rather, because these functions are split into two modalities by commissurotomy, consciousness must be primarily associated with neocortical activities.
Consider, however, the proposition that though localized brain processes are certainly the necessary physical substrate of consciousness awareness, they are not also the sufficient physical substrate. Physical processes other than that of the nervous system are involved in mediation of conscious experience. As A. E. Fessard once said, “They [cybernetic schemata] aid us to escape at one and the same time both the naivetes of strict localizationalism and the confusions of integral holisticism” [as quoted in 157,
p. 120]. This statement provides a clue. The localization and fusion process is instrumental in manifesting processes of consciousness.
As a result of research [157-160] on residual tension patterns of the voluntary muscle system in relation to mental functions, which involved simultaneous electrophysiological measurements of various parameters and subjective reports by individuals highly trained in autosensory observation, Edmund Jacobson was able to demonstrate:
… that in mental activities of all forms there are no closed physiological circuits in the brain. Our evidence suggests that at the moment of occurrence of any form of mental activity, the physiological circuits include not only associative, afferent and efferent brain neurones, axons, and dendrites, but also afferent and efferent nerve activities and peripheral activities chiefly in the form of muscle contraction. The contraction turns out to be specifically patterned in each such moment. [157, p. 95.]
Through the study of simultaneous electroencephalographic and electromyographic recordings, Jacobson was led to question the commonly held idea that brain action precedes muscular contraction. In the case of the intact organism, where no electrical stimulation of the brain is used, he concluded that, “Central nervous regions do not prevail over an allegedly subordinate peripheral region” [157, p. 110], but, rather, that during mental activity the central nervous system and neuromuscular system operate simultaneously through a mechanism of “reciprocating activation”. Regarding the question of mind-body interaction, he says:
According to the present argument, the control of mental activity and of behavior has never been experimentally identified and will not be recorded in any portion of the organism, because it resides in the integrated totality. [157, p. 91.]
But if it is no longer to be thought that the brain alone controls behavior, in what context are the processes of activation and attention to be understood? The proper context is that established by spontaneous localization and fusion. Consider the digital data system (the sum total of molecular and cellular processes constituting the nervous system) to be a decomposition of the information content of the quantum wave phenomena and associated frequency regime (i.e., the analog system). Generally, there is a tendency to think of the molecules as pre-existing and generating the wave phenomena. This is because there is greater familiarity with molecular processes, because they can be directly imaged and structurally modeled, and because they were discovered first. But, actually, the quantum wave processes are more fundamental and a shift in the point of view generates a new perspective.
The processing of information is handled in a multivalued fashion by the quantum-wave analog-type data system, while the digital data system, essentially, is a localized, automatized input-output system-control mode for the information content in its decomposed form. The decomposition process is the process of spontaneous localization, wherein multivalued information is decomposed into a hierarchically-ordered, single-valued, localized information system. The inverse of this is the recomposition process or spontaneous fusion. The information input undergoes spontaneous fusion before being processed in a multivalued, nonlocal, linear-time independent fashion, and then spontaneous localization before being outputted into conscious awareness in the single-valued, localized, linear-time dependent modality. Just as the integrating functions of the reticular activating and centrencephalic systems (tonic and phasic activation) have been hypothesized to mediate possible conflictual functioning between cortical and subcortical mechanisms [7, 133, 137, 145-148,
161-164], so we hypothesize that, via the localization and fusion process, a “relative state” activation of the brain mediates possible conflictual functioning between central and peripheral systems, and, hence, contributes significantly to functional integration of the organism as a whole and to the high levels of efficiency in psychosomatic interaction pre-requisite to heterostasis.
A Wave STATEMENT for an Autopoietic Process
What stereochemical processes are associated with spontaneous localization and fusion? The hypothesis we shall adopt is that neuronal and glial DNA are instrumental in the mediation, respectively, of the processes of localization and fusion. To discuss this concept in some detail, let us return to the dynamic model, proposed by the present authors, describing DNA as superconductant [54]. Superconductivity of the DNA molecule is described as resulting from scale interaction processes mediated by a radiation-induced, harmonic oscillation of a parcel of p-electrons (a domain structure) between the reference level and critical temperatures. The fundamental frequency of the domain structure is established as being inversely related to the minimum time for spontaneous localization. This minimum time determines the parcel�s period of transit between To«Tc. A change in this period signifies that the DNA molecule is receiving additional radiation from the environment which necessitates a different time rate of change of bonding. This change in the DNA molecule�s activity is communicated within the molecule as well as into the DNA environment by coherent waves.
To develop these coherent wave equations, we chose to designate the bonded state representative of a mass-momentum balance within the DNA molecule by the symbol F. The unbonded state, typified by an accelerating p-electron parcel, was designated by the symbol X . In order that the temporal period (no-1) for parcel transit To«Tc resulting from radiation impingement may be transduced from that of a purely temporal mode to one of spatial representation, it is necessary to define three limiting parameters:
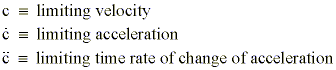
The composite wave statement is:
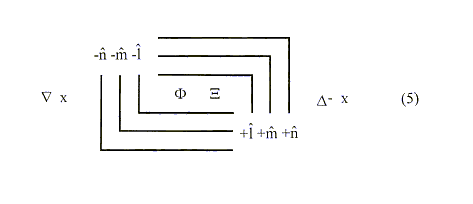
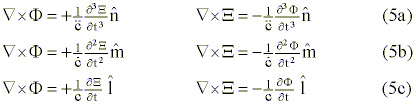
where each of the foregoing critical parameters
(Ñ x F, Ñ x X) into that of a temporal representation given by the above partial derivatives taken with respect to time.
Equations (5a), (5b), and (5c) treat the DNA molecule as a self-referential (self-reentrant), open, and fully-isolated information system, respectively. The collapse of the wave function [j (F, X)] is represented by reading the set of equations from (5a) through (5b) to (5c). The wave function�s recomposition requires reading the same set in the reverse direction. The appendix provides more background regarding this formulation.
DNA-Mediated Quantal Localization and Fusion in Cortical Function
and Hyden�s Molecular Theory of Memory
On the basis of this model of superconductant DNA, it is hypothesized that the macroquantum localization process is recorded as phasic cortical activation by a free-energy cascade through the nested hierarchy of organizational levels in the cortex. The interference patterns in the quantum frequency domain decompose into a single-valued representation through resonance exchange initiated on the quantum level and proceeding through the molecular and cellular levels. As interference patterns in the frequency domain change and impinge upon the p-electron parcels associated with the structure of DNA molecules (within the soma of the neurons comprising the cortical modules), these parcels undergo a sequence of critical-state radiation exchanges. Variations in the referencing frequency domain serve to amplify or damp the fundamental frequency of a given parcel of p-electrons. Specific temporal patternings of these radiation exchanges (distinguished by very small time differences) trigger frequency responses in the ensemble of p-electron parcels associated with a given DNA molecule. These frequency modulations are instrumental in governing the helix-coil transitions of the molecule, where the inverse of a specific frequency determines the minimum time for spontaneous localization. As frequency variations occur, there are correlated variations in the respective minimum times, which, in turn, determine the time rate of change of bonding. Given existing knowledge of RNA synthesis [166], it appears likely that a given minimum time for spontaneous localization is correlated with a given RNA molecule such that when a given frequency is attained it serves as a start signal for RNA polymerase to attach to the DNA molecule and begin synthesis. The given specific RNA molecule then synthesizes specific proteins, which, in turn, initiate synaptic transmitter production. As this localization process transpires in a multiplicity of neurons within in given cortical module, a linear-time dependent pattern of activation is established and chemically specified (see Figure 2). Similarly, as a multiplicity of modules in a region of cortex are activated in this fashion, linear-time dependent patterns of module interaction are established. Thus, through the process of spontaneous localization, spatiotemporal patterns of cortical activity occur in response to specific patterns in the quantum-level referencing frequency domain.
These ideas are in many respects similar to H. Hyden�s molecular theory of memory [10, 167-172], which is summarized in the following statement:
I would suggest that the time pattern of frequencies set up in the neurons involved leads to a release of repressed regions of chromosomal DNA. This leads to a production of highly specific DNA-copied RNA. It its turn, the RNA synthesis occurring on the demand of the situation gives, as an end product in the neural soma, specific proteins. The presence of these proteins or, at later stages, their rate of production, leads to an activation of the transmitter substance. The next time the same modulated frequencies enter, these specific proteins answer with a rapid reaction leading to an activation of the transmitter substance. In analogy to the mechanism of antigen-antibody reaction, the specific proteins react to the same modulated frequencies which first led to the release of the chromosomal activity, the synthesis of the DNA-dependent RNA, and their own formation. [171, p. 33.]The process of spontaneous fusion, on the other hand, is hypothesized to be mediated by glial DNA because the research of R. Becker [82] has suggested that the perineural cell system (Schwann cells peripherally and glial cells centrally) is fundamentally related to the analog data system. I. Tasaki and J. J. Chang have described analog type electrical activity as accompanying glial cell function [82, 173]. It is proposed, then, that the continuously changing electrical activity in the glial cells sets up highly interactive standing wave fronts which form holographic representations as a result of glial DNA serving as the coherent wave source required to generate the hologram. As the superconductant DNA model proposes that the coherent waves are generated by the DNA molecule in response to scale interaction, the holographic pattern represents the systematic integration (i.e., the “relative state”) of the organism.
Relative-state activation transpires as a result of spontaneous localization and fusion, mediated by the neural and perineural tissues. Increases in phasic activation (ergotropic) correspond to increased localization of information, while decreases in phasic activation (trophotropic) correspond to increased fusion of information. Specificity and integration of growth and repair processes require efficient information localization and fusion in the system hierarchy. As the efficiency of this process increases, the ground state (or characteristic base line, with its associated baud rate of consciousness) of tonic activation is shifted from that necessary to maintain homeostasis toward that of heterostasis.
Autogenic shift has been hypothesized to involve a relative movement from ergotropically- to trophotropically-oriented mechanisms, involving alterations in reticulo-cortical and thalamo-cortical activity [128, pp. 3-6]. Given the present context, this requires a prior state of imbalance between localization and fusion recorded in effects of neuronal material possessing pathofunctional potency. Autogenic shift, thus, represents an attempt of the organism to normalize this prior imbalance through neutralization of pathogenic neuronal material. The improved efficiency is reflected in a shift of the ground state of tonic activation.
Quantum Measurement, Autogenic Discharge,
and Biological Information Exchange at Critical States
On basis of the above discussion, it now appears possible to formulate a hypothesis concerning the fundamental role of autogenic discharge activity in organization of a biological process. The hypothesis we propose is that the autogenic discharge is identical to the process of spontaneous localization in quantum measurement (which occurs in all perception and proprioception), wherein multivalued information in the referencing frequency domain is decomposed into a single-valued representation and communicated throughout the nested hierarchy of organismic organizational levels. In other words, the clinical and subclinical autogenic discharge is identical to collapse of the state function of the organism. It is, therefore, hypothesized that during autogenic standard exercises the self-observation involved in passive concentration upon prescribed psychophysiologic structures is a special case of the quantum mechanical observer-state object-system interaction. Luneburg�s experimental findings concerning localization in visual space as learned behavior very strongly support this notion of spontaneous localization in quantum measurement as autogenic discharge. If ability to localize (in visual space) the needle�s position on the gauge of a measuring instrument depends on consensuated (a collective occasion of experience) psychological factors, as Luneburg�s findings clearly imply, then this (cooperative) localization of the needle�s position on the gauge must take place in the brains of participants or in association with some other substrates of conscious awareness.
According to this hypothesis concerning the quantum mechanical basis of self-observation, autogenic discharges are occurring continually at a subclinical level of intensity. These discharges are a fundamental requirement for maintenance of growth and repair processes, in that they return altered values of biological clocks and immunological signifiers to the normative. The clinical phenomena observed during therapeutic application of the methods of autogenic therapy (to include those associated with use of organ-specific training formulas) are the result of a prior condition in the patient which has undermined functional integrity of the mechanisms governing the localization and fusion process.
In order to examine this perspective more specifically, consider the following statement by S. Bogoch made in The Biochemistry of Memory:
The mechanisms by which organ-specific substances live in immunological isolation side by side in the same organism, and the breakdown of this isolation which leads to �autoimmune disease�, are fascinating and still poorly understood aspects of the handling of immunological information that may have parallels in the compartmentalization of information in the nervous system and its breakdown. [167, p. 13.]In terms of the present macroquantum description, this isolation is attained as a result of the DNA molecules of a given neural compartment being receptive only to a narrow frequency window of the multitude of frequencies inherent in the referencing frequency domain of the organism. As the multivalued information is localized and transformed into a single-valued representation, it becomes compartmentalized on the basis of frequency windows. The following quotation from A. S. Presman bears directly upon this question:
If the described effects are considered from the standpoint of the concept of diverse EmF interactions within the organism and its interactions with environmental EmFs, then we can sketch a picture which is very convincing in its simplicity and consistency. In fact, we can picture the organism provided with diverse interconnections of such kind (in addition, of course, to the known diverse neurohumoral connections), differentiated as regards their specific “working” frequencies, intensity ranges, and method of coding. Such interconnections may underlie not only the interactions between cells, but also specific interactions between macromolecules: enzyme and substrate, antigen and antibody, DNA and RNA. Similar interconnections may be responsible for the control of protein synthesis. In a recently proposed hypothesis regarding such control (New Biological Effects of R-F, 1959) DNA molecules are regarded as generators of radio-frequency signals, RNA molecules as amplifiers, and enzymes and amino acids as effectors of signals coded in various regions of the spectrum; the cell wall is believed to be a noise filter. [90, p. 243.]Integrity of the frequency windows underlying immunological isolation and neurological compartmentalization is dependent upon viability of electron transport chains and unimpaired functioning of p-electron parcels in macromolecular processes underlying cell metabolism. As described in the superconductant DNA model, these are scale-interactive critical state processes by which the relative state of the organism is encoded, communicated, and maintained.
It is the role of clinical and subclinical autogenic discharge activity to maintain integrity of these processes. But how would the autogenic discharge maintain functional integrity if there were a culturally reinforced chronic resistance to the multivalued functional modality? In view of the present macroquantum description of biological information exchange and Luneburg�s findings regarding localization of objects in binocular visual space, it appears further investigation of this question is in order. It has been implicitly felt that, if the “classical limit” were swept away and full applicability of quantum principles to everyday events established, the world would appear a phantasmagoria where objects would not hold their place and hallucination would become the norm. Is this, indeed, the case? Fear that it is has been part and parcel of the current crisis in psychiatry and medicine. Or is it that this implicit assumption is the result of prescriptive enculturation, where dissociative experience is branded inevitably pathological? Is it possible that the widespread culturally reinforced fear of dissociation has predisposed the quantum theoretician in his analysis of the meaning of the equations of quantum mechanics? The tendency not to take the formalism of quantum mechanics at face value, and to generate metaphysical interpretations instead, may be necessitated by a culturally transmitted avoidance of dissociative experience. Were it not for this enculturated negative assumption, some types of dissociative experience would be healthy, even therapeutic. The aforementioned “functional disintegration” or “splitting up for release” of neuronal material during autogenic abreaction is an indication that dissociative experience can have therapeutic value. The purely negative assumption regarding dissociation clearly constitutes a particularly potent variety of the “thematic evasion” type of “neutralization antagonizing forms of resistance” (NAFR) [3, pp. 14-16] which minimize therapeutic value of the abreaction process. New insight into the quantum measurement problem requires simultaneous new insight into the nature of sensory experience (particularly visual phenomena: see W. Luthe, “The Clinical Significance of Visual Phenomena During the Autogenic State” for background) [128] during “schizophrenic breaks” and “hysterically induced” states of dissociation.
The prior condition in the patient which undermines functional integrity of mechanisms governing localization and fusion is, at least in part, a direct result of culturally reinforced chronic resistance to the multivalued functional modality. When NAFR become chronic as a result of prescriptive enculturation there is a disturbance in the efficiency of linear-time independent macroquantum oscillation between the composite and decomposed modalities. This is to say that the macroquantum information exchange process is disrupted in such a manner as minimize information flow or to cause inaccuracies in the information transmitted. When these resistances are lessened or removed, as, for example, during therapeutic application of the methods of autogenic therapy, there is massive unloading manifest as clinically observable discharge phenomena. These phenomena are representative of an attempt by the system to re-establish the functional integrity of its information transport mechanisms.
It would, therefore, be appropriate to suggest that the structure of the unenculturated organism, and the bare universe (Realitat as opposed to Wirklichkeit) of which it is a part, is fundamentally multivalued and composed of parts that are only quasi-separable through the macroquantum localization process. Prescriptive enculturated programming of cognitive development attempts to impose a purely single-valued, localized linear-time dependent functional modality to exclusion of the underlying multivalued, nonlocal, linear-time independent structure of both perceptual mechanisms and metabolic processes. This imposition of prescriptive enculturation disrupts functional viability of integrative mechanisms and immune system, thus contributing significantly to pathogenesis of degenerative disease and schizophrenia. These diseases constitute a functional disorientation to the quantum mechanical basis of observer state and organic system function. Autogenic discharges are an attempt of the organism to re-establish functional coherence of its fundamental integrative mechanisms. Bare perception, spontaneous visualization, and many types of altered states of consciousness are inherently therapeutic because they facilitate the process of re-instating functional viability of the multivalued mechanisms underlying functional integration.
(View Musings, Reflections, and Sketches as this Paper Gestated.)
(For further information see: “Four Conversations”.)
- W. Luthe. Autogenic Therapy, 4. New York: Grune and Stratton, 1970.
- W. Luthe. Autogenic Therapy, 5. New York: Grune and Stratton, 1970.
- W. Luthe. Autogenic Therapy, 6. New York: Grune and Stratton, 1973.
- W. Luthe. Autogenic Therapy, 2. New York: Grune and Stratton, 1969.
- W. Luthe. Autogenic Therapy, 3. New York: Grune and Stratton, 1969.
- J. F. Delafresnjaye (ed.). Brain Mechanisms and Consciousness. Springfield, Ill.: C. C. Thomas, 1954.
- W. Penfield and H. Jasper. Epilepsy and the Functional Anatomy of the Human Brain. Boston: Little, Brown and Company, 1954.
- W. Penfield and L. Roberts. Speech and Brain Mechanisms. Princeton: Princeton U. Press, 1959.
- J. C. Eccles (ed.). Brain and Conscious Experience. New York: Springer-Verlag, 1966.
- J. C. Eccles. Facing Reality. New York: Springer-Verlag, 1970.
- J. C. Eccles, “The Physiology and Physics of the Free Will Problem”, in
S. L. Mintz and S. M. Widmayer (eds.). Progress in the Neurosciences and Related Fields. New York: Plenum Press, 1974. - G. Globus, G. Maxwell and I. Savodnik (eds.). Consciousness and the Brain: A Scientific and Philosophical Inquiry. New York: Plenum Press, 1976.
- R. W. Sperry, “Neurology and the Mind-Brain Problem”, in R. L. Isaason (ed.). Basic Readings in Neuropsychology. New York: Harper and Row, 1964.
- R. W. Sperry, “A Modified Concept of Consciousness”, Psychological Review, 76:6, pp. 532-536, 1969.
- R. W. Sperry, “In Search of Psyche”, in F. G. Worden, J. P. Swazey and G. Adelman (eds). The Neurosciences: Paths of Discovery. Cambridge: M.I.T. Press, 1975.
- W. Penfield. The Mystery of the Mind. Princeton: Princeton U. Press, 1975.
- L. M. Beynam, “Quantum Physics and Paranormal Events”, in J. White and
S. Krippner (eds.). Future Science. New York: Anchor, 1977. - L. N. Cooper and D, Van Vechten, “On Interpretation of Measurement in the Quantum Theory”, American Journal of Physics, 37:12, pp. 1212-1220, 1969.
- B. S. De Witt, “Quantum Mechanics and Reality”, Physics Today, 23:9,
pp. 30-35, 1970. - C. A. Hooker, “Concerning Einstein�s Podolsky�s and Rosen�s Objection to Quantum Theory”, American Journal of Physics, 38:7, pp. 851-857, 1970.
- E. H. Walker, “The Nature of Consciousness”, Mathematical Biosciences, 7:1/2, pp. 131-178, 1970.
- E. H. Walker, “Consciousness as a Hidden Variable”, Physics Today, 24:4,
p. 39, 1971. - E. H. Walker, “Foundations of Paraphysical and Parapsychological Phenomena”, in Laura Oteri (ed.). Quantum Physics and Parapsychology. New York: Parapsychology Foundation, 1975.
- E. P. Wigner, “Remarks on the Mind-Body Question”, in I. J. Good (ed.).
The Scientist Speculates. London: William Heinemann, 1961. - E. P. Wigner. Symmetries and Reflections. Bloomington: Indiana U. Press, 1967.
- E. P. Wigner, “The Place of Consciousness in Modern Physics”, C. Muses and A. M. Young (eds.). Consciousness and Reality. New York: Avon, 1969.
- W. Luthe. Autogenic Therapy, 1. New York: Grune and Stratton, 1969.
- T. Bastin (ed.). Quantum Theory and Beyond. Cambridge: Cambridge U. Press, 1971.
- F. J. Belinfante. A Survey of Hidden Variable Theories. New York: Pergamon, 1973.
- D. Bohm, “A Suggested Interpretation of the Quantum Theory in terms of �Hidden� Variables”, Physical Review, 85:2, pp. 166-179, 1952 and ibid.,
pp. 180-193. - M. Bunge (ed.). Quantum Theory and Reality. New York: Springer-Verlag, 1967.
- B. S. De Witt and N. Graham (eds.). The Many-Worlds Interpretation of Quantum Mechanics. Princeton: Princeton U. Press, 1973.
- B. D�Espagnat. Conceptual Foundations of Quantum Mechanics, 2nd ed. Menlo Park: Benjamin-Cummings, 1976.
- H. P. Stapp, “S-Matrix Interpretation of Quantum Theory”, Physical Review D, 3:6, pp. 1303-1320., 1970.
- J. von Neumann. Mathematical Foundations of Quantum Mechanics. Princeton: Princeton U. Press, 1955.
- M. Siegler and H. Osmond. Models of Madness, Models of Medicine.
New York: Harper and Row, 1974. - H. Everett, III, “�Relative State� Formulation of Quantum Mechanics”, Reviews of Modern Physics, 29:3, pp. 454-462, 1957.
- H. Everett, III, “The Theory of the Universal Wave Function”, B. S. De Witt and N. Graham (eds.). The Many-Worlds Interpretation of Quantum Mechanics. Princeton: Princeton U. Press, 1973.
- P. J. Werbos, “Experiments on the Reinterpretation of Quantum Mechanics: Corrections and New Ideas”, IL Nuovo Cimento, 37B:1, pp. 24-33, 1977.
- B. S. De Witt, “The Many-Universes Interpretation of Quantum Mechanics”, B. S. De Witt and N. Graham (eds.). The Many-Worlds Interpretation of Quantum Mechanics. Princeton: Princeton U. Press, 1973.
- B. J. Hiley, “Foundations of Quantum Mechanics”, Contemporary Physics, 18:4, pp. 411-414, 1977.
- J. Piaget, “Principle Factors Determining Intellectual Evolution From Childhood to Adult Life”, in D. Rappaport (ed.). Organization and Pathology of Thought. New York: Columbia U. Press, 1951.
- H. G. Furth. Thinking Without Language. New York: Free Press, 1966.
- R. K. Luneburg, “The Metric of Binocular Visual Space”, Journal of the Optical Society of America, 40:10, pp. 627-642, 1950.
- A. A. Blank, “The Luneburg Theory of Binocular Visual Space”, Journal of the Optical Society of America, 43:9, pp. 717-727, 1953.
- A. Baracca, D. J. Bohm, B. J. Hiley and A. E. G. Stuart, “On Some New Notions Concerning Locality and Nonlocality in the Quantum Theory”,
IL Nuovo Cimento, 28B:2, pp. 453-465, 1975. - D. Bohm, “Quantum Theory as an Indication of a New Order in Physics.
“Part A. The Development of New Orders as Shown Through the History of Physics”, Foundations of Physics, 1:4, pp. 359-381, 1971. And “Part B. Implicate and Explicate Order in Physical Law”, ibid., 3:2, pp. 139-168, 1973. - D. J. Bohm and B. J. Hiley, “On the Intuitive Understanding of Nonlocality as Implied by Quantum Theory”, Foundations of Physics, 5:1, pp. 93-109, 1975.
- K. Pribram. Languages of the Brain. Englewood Cliffs: Prentice-Hall, 1971.
- K. Pribram, “Problems Concerning the Structure of Consciousness”, in
G. Globus, G. Maxwell and I. Savodnik (eds.). Consciousness and the Brain: A Scientific and Philosophical Inquiry. New York: Plenum Press, 1976. - K. Pribram, “Some Comments on the Nature of the Perceived Universe”,
R. Shaw and J. Bransford (eds.). Perceiving, Acting, and Knowing. Hillsdale: Lawrence Erlbaum, 1977. - D. Goleman, “Holographic Memory: Karl Pribram Interviewed”, Psychology Today, 12;9, pp. 70-84, Feb., 1979.
- M. Ferguson, “Primary Reality May Be Frequency Realm, Theory Says”, Brain-Mind Bulletin, 2:16, pp. 1-4, July 4, 1977.
- D. A. Paine and W. L. Pensinger, “A Dynamical Theory Describing Superconductant DNA”, International Journal of Quantum Chemistry, 15:3, pp. 333-341, 1979.
- J. L. Synge, “The Electrodynamic Double Helix”, J. R. Klauder (ed.). Magic Without Magic: John Archibald Wheeler. San Francisco: W. H. Freeman, 1972.
- A. H. Klopf. Brain Function and Adaptive Systems - A Heterostatic Theory. L. G. Hanscom Field, Bedford, Mass: Air Force Cambridge Research Laboratories Special Report No. 133 (AFCRL-72-0164 and DDC Report No. AD 742259), 1972.
- W. L. Pensinger and D. A. Paine, “Deautomatization and the Autogenic Discharge”, Journal of Altered States of Consciousness, 3:4, pp. 325-335, 1977-78.
- H. S. Burr, “An Electrodynamic Theory of Development Suggested by Studies of Proliferation Rates in the Brain of Amblystoma”, Journal of Comparative Neurology, 56:2, pp. 347-371, 1932.
- H. S. Burr and C. T. Lane, “Electrical Characteristics of Living Systems”,
Yale Journal of Biology and Medicine, 8, pp. 31-35, 1935. - H. S. Burr and C. I. Hovland, “Potential Gradients in the Chick”, Yale Journal of Biology and Medicine, 9, pp. 247-258, 1937.
- H. S. Burr, “Electrical Correlates of Pure and Hybrid Strains of Corn”, Proceedings of the National Academy of Sciences, 29:6, pp. 163-166, 1943.
- O. E. Nelson and H. S. Burr, “Growth Correlates of Electromotive Forces in Maise Seeds”, Proceedings of the National Academy of Sciences, 32:4,
pp. 73-84, 1946. - H. S. Burr and W. Seifrietz, “Response of the Slime Mold to Electric Stimulus”, Science, 122: 3178, pp. 1020-1021, 1955.
- H. S. Burr and F. S. C. Northrop, “The Electro-Dynamic Theory of Life”, Quarterly Review of Biology, 10:3, pp. 322-333, 1935.
- H. S. Burr, “Biologic Organization and the Cancer Problem”, Yale Journal of Biology and Medicine, 12, pp. 277-282, 1940.
- H. S. Burr, S. C. Harvey and M. Traffel, “Bioelectric Correlates of Wound Healing”, Yale Journal of Biology and Medicine, 11, pp. 104-107, 1938.
- L. Langman and H. S. Burr, “Electro-Magnetic Studies in Women with Malignancy of Cervix Uteri”, Science, 105: 2721, pp. 209-210, 1947.
- T. S. England, “Dielectric Properties of the Human Body for Wavelengths in the 1-10 cm Range”, Nature, 166:4220, pp. 480-481, 1950.
- T. S. England and N. A. Sharpless, “Dielectric Properties of the Human Body in the Microwave Region of the Spectrum”, Nature, 163:4143, pp. 487-488, 1949.
- H. P. Schwan, “Electrical Properties of Blood at Ultrahigh Frequencies”, American Journal of Physical Medicine, 33:3, pp. 144-152, 1953.
- H. P. Schwan and G. M. Piersol, “The Absorption of Electromagnetic Energy in Body Tissues. Part I. Biological Aspects”, American Journal of Physical Medicine, 33:6, pp. 371-404, 1954.
- H. Cook, “Dielectric Behavior of Human Blood at Microwave Frequencies”, Nature, 168:4267, pp. 247-248, 1951.
- R. O. Becker and R. M. Brown, “Photoelectric Effects in Human Bone”, Nature, 206:4991, pp. 1325-1328, 1965.
- H. Athenstaedt, “Pyroelectric and Piezoelectric Properties of Vertebrates”, Annals of the New York Academy of Sciences, 238, pp. 68-93, 1974.
- S. B. Lang, “Pyroelectric Effect in Bone and Tendon”, Nature, 212:5063,
pp. 1325-1328, 1965. - E. Fukada and I. Yasuda, “Piezoelectric Effects in Collagen”, Japanese Journal of Applied Physics, 3:2, pp. 117-121, 1964.
- M. Barnothy (ed.). Biological Effects of Magnetic Fields. New York:
Plenum Press, 1964. - C. A. L. Bassett, R. J. Pawluk and R. O. Becker, “Effects of Electric Currents on Bone In Vivo”, Nature, 204:4959, pp. 652-654, 1964.
- R. O. Becker, “The Bioelectric Factors in Amphibian Limb Regeneration”, Journal of Bone and Joint Surgery, 43A:5, pp. 643-656, 1961.
- R. O. Becker, “Relationship of Geomagnetic Environment to Human Biology”, New York State Journal of Medicine, 63:15, pp. 2215-2219, 1963.
- R. O. Becker, “Stimulation of Partial Limb Regeneration in Rats”, Nature, 235:5332, pp. 109-11, 1972.
- R. O. Becker, “The Basic Biological Data Transmission and Control System Influenced by Electrical Forces”, Annals of the New York Academy of Sciences, 238, pp. 236-241, 1974.
- R. O. Becker, C. H. Bachman and F. Friedman, “The Direct Current Control System: A Link Between Environment and Organism”, New York State Journal of Medicine, 62:8, pp. 1169-1176, 1962.
- R. O. Becker and J. A. Spadaro, “Electrical Stimulation of Partial Limb Regeneration in Mammals”, Bulletin of the New York Academy of Medicine, 48:4, pp. 627-641, 1972.
- G. V. B. Cochran, “Experimental Methods for Stimulation of Bone Healing by Means of Electrical Energy”, Bulletin of the New York Academy of Medicine, 48:7, pp. 899-911, 1972.
- H. J. Hambury, J. Watson, A. Sivyer and D. J. B. Ashley, “Effect of Microamp Electrical Currents on Bone In Vivo and its Measurement Using Strontium-85 Uptake”, Nature, 231:5299, pp. 190-191, 1971.
- C. E. Humphrey and E. H. Seal, “Biophysical Approach Toward Tumor Regression in Mice”, Science, 130:3372, pp. 388-389, 1959.
- L. S. Lavine, I. Lustrin, M. H. Shamos and M. L. Moss, “The Influence of Electric Current on Bone Regeneration In Vivo”, Acta Orthopedica Scandinavica, 42:4, pp. 305-314, 1971.
- C. Romero-Sierra and J. A. Tanner, “Biological Effects of Nonionizing Radiation: An Outline of Fundamental Laws”, Annals of the New York Academy of Sciences, 238, pp. 263-272, 1974.
- A. S. Pressman. Electromagnetic Fields and Life. New York: Plenum Press, 1970.
- A. R. Liboff and R. A. Rinaldi (eds.), “Electrically Mediated Growth Mechanisms in Living Systems” (volume title), Annals of the New York Academy of Sciences, 238, 1974.
- D. Brenner, S. J. Williamson and L. Kaufman, “Visually Evoked Magnetic Fields of the Human Brain”, Science, 190:4213, pp. 480-482, 1975.
- D. Cohen, “Magnetic Fields Around the Torso: Production by Electrical Activity of the Human Heart”, Science, 156:3775, pp. 652-654, 1967.
- D. Cohen, “Magnetoencephalography: Evidence of Magnetic Fields Produced by Alpha-Rhythm Currents”, Science, 161:3843, pp. 784-786, 1968.
- D. Cohen, “Magnetoencephalography: Detection of the Brain�s Electrical Activity with a Superconducting Magnetometer”, Science, 175:4022,
pp. 664-666, 1972. - H. S. Burr and A. Mauro, “Electro-Static Fields of the Sciatic Nerve in the Frog”, Yale Journal of Biology and Medicine, 21, pp. 455-462, 1949.
- A. Fraser and A. H. Frey, “Electromagnetic Emission at Micron Wavelengths from Active Nerves”, Biophysical Journal, 8:6, pp. 731-734, 1968.
- K. H. Pribram, “The Neurophysiology of Remembering”, Scientific American, 220:1, pp. 73-86, 1969.
- J. Aschoff, “Exogenous and Endogenous Components in Circadian Rhythms”, Cold Spring Harbor Symposia on Quantitative Biology, 25, pp. 11-28, 1960.
- F. A. Brown, “Living Clocks”, Science, 130:3388, pp. 1535-1544, 1959.
- F. A. Brown, “The Clocks Timing Biological Rhythms”, American Scientist, 60:6, pp. 756-766, 1972.
- F. A. Brown, “Response to Pervasive Geophysical Factors and the Biological Clock Problem”, Cold Spring Harbor Symposia on Quantitative Biology, 25, pp. 57-71, 1960.
- F. A. Brown, “Extrinsic Rhythmicality: A Reference Frame for Biological Rhythms Under So-Called Constant Conditions”, Annals of the New York Academy of Sciences, 98, pp. 775-787, 1962.
- F. Halberg, “Temporal Coordination of Physiological Function“, Cold Spring Harbor Symposia on Quantitative Biology, 25, pp. 289-310, 1960.
- R. Foster. Organic Charge Transfer Complexes. London: Academic Press, 1969.
- M. A. Slifkin. Charge Transfer Interactions of Biomolecules. London: Academic Press, 1971.
- R. S. Mulliken, “Molecular Compounds and Their Spectra, II”, Journal of the American Chemical Society, 74:3, pp. 811-824, 1952.
- R. S. Mulliken, “Molecular Compounds and Their Spectra, III, The Interaction of Electron Donors and Acceptors”, Journal of Physical Chemistry, 56:7,
pp. 801-822, 1952. - A. Szent Györgyi. Bioenergetics. New York: Academic Press, 1957.
- A. Szent Györgyi. Introduction to a Submolecular Biology. New York: Academic Press, 1960.
- A. Szent Györgyi. Bioelectronics. New York: Academic Press, 1968.
- A. Szent Györgyi. The Living State. New York: Academic Press, 1972.
- A. Szent Györgyi. Electronic Biology and Cancer. New York:
Marcel Dekker, 1976. (Current comment by Derek Dillon: Whenever one sees a two-time Nobel laureate unable to get his publisher to publish the scientific monograph which is the without-logical-leap extension of his prior monographs published by this same publisher over the previous two decades, one realizes that the scientific field in question has become so deeply corrupted that nothing one reads in the scientific journals can be taken at face value.) - S. Freud, “Formulations Regarding the Two Principles of Mental Functioning”, in D. Rapaport (ed.). Organization and Pathology of Thought. New York: Columbia U. Press, 1951.
- D. Rapaport, “Toward a Theory of Thinking”, in D. Rapaport (ed.). Organization and Pathology of Thought. New York: Columbia U. Press, 1951.
- H. Hartmann. Ego Psychology and the Problem of Adaptation. New York: International Universities Press, 1958.
- D. Rapaport, “The Theory of Attention Cathexis”, in M. M. Gill (ed.). The Collected Papers of David Rapaport. New York: Basic Books, 1967.
- M. M. Gill and M. Brenman. Hypnosis and Related States. New York: International Universities Press, 1959.
- A. F. Deikman, “Deautomatization and the Mystic Experience”, Psychiatry, 29:4, pp. 324-338, 1966.
- L. E. Marks, “On Colored-Hearing Synesthesia: Cross-Modal Translations of Sensory Dimensions”, Psychological Bulletin, 83:3, pp. 303-331, 1975.
- E. R. Jaensch. Eidetic Imagery. New York: Harcourt, Brace, and Company, 1930.
- L. J. Ravitz, “Electromagnetic Correlates of the Hypnotic State”, Science, 112:2908, pp. 341-342, 1950.
- L. J. Ravitz, “Application of Electro-Dynamic Field Theory in Biology, Psychiatry, Medicine, and Hypnosis, I, General Survey”, American Journal of Clinical Hypnosis, 1:4, pp. 135-150, 1959.
- L. J. Ravitz, “Standing Potential Correlates of Hypnosis and Narcosis”, Archives of Neurology and Psychiatry, 65:4, pp. 413-436, 1951.
- L. J. Ravitz, “History, Measurement and Applicability of Periodic Changes in the Electro-Magnetic Field in Health and Disease”, Annals of the New York Academy of Sciences, 98, pp. 1144-1201, 1962.
- L. J. Ravitz, “Electro-Magnetic Field Monitoring of Changing State-Function, Including Hypnotic States”, Journal of the American Society of Psychosomatic Dentistry and Medicine, 17:4, pp. 119-127, 1970.
- L. J. Ravitz, “Studies of Man in the Life Field”, Main Currents in Modern Thought, 19:1, pp. 13-23, 1962.
- W. Luthe, A. Jus, and P. Geissmann, “Autogenic State and Autogenic Shift: Psychophysiologic and Neurophysiologic Aspects”, in W. Luthe (ed.). Autogenic Training. New York: Grune and Stratton, 1965.
- K. G. Wilson, “Problems in Physics With Many Scales of Length”, Scientific American, 241:2, pp. 158-179, 1979.
- H. Haken and M. Wagner (eds.). Cooperative Phenomena. New York: Springer-Verlag, 1973.
- G. Ungar, “Is the Brain a Chemical Computer?”, in S. L. Mintz and S. M. Widmayer (ed.). Progress in the Neurosciences and Related Fields.
New York: Plenum Press, 1974. - J. D. French, “The Reticular Formation”, Scientific American, 196:5,
pp. 54-60, 1957. - H. H. Jasper, “Diffuse Projection Systems: The Integrative Action of the Thalamic Reticular System”, Electroencephalography and Clinical Neuro-physiology, 1:4, pp. 405-420, 1949.
- H. W. Magoun, “The Ascending Reticular Activating System”, Research Publications: Association for Research in Nervous and Mental Disease, 30:4, pp. 480-492, 1952.
- G. Moruzzi and H. W. Magoun, “Brain Stem Reticular Formation and Activation of the EEG”, Electroencephalography and Clinical Neurophysiology, 1:4, pp. 455-473, 1949.
- J. W. Papez, “Central Reticular Path to Intralaminar and Reticular Nuclei of Thalamus for Activating EEG Related to Consciousness”, Electroencephalo-graphy and Clinical Neurophysiology, 8:1, pp. 117-128, 1956.
- A. B. Rathballer, “Studies on the Adrenaline-Sensitive Component of the Reticular Activating System”, Electroencephalography and Clinical Neuro-physiology, 8:4, pp. 602-621, 1956.
- T. E. Starzl and H. W. Magoun, “Organization of the Diffuse Thalamic Projection System”, Journal of Neurophysiology, 14:2, pp. 133-146, 1951.
- J. C. Eccles. The Neurophysiological Basis of Mind: Principles of Neurophysiology. Oxford: Claredon Press, 1953.
- J. C. Eccles. The Physiology of Synapses. New York: Springer-Verlag, 1964.
- J. C. Eccles, M. Ito and J. Szentagothai. The Cerebellum as a Neuronal Machine. New York: Springer-Verlag, 1967.
- V. B. Mountcastle, “Modality and Topographic Properties of Single Neurones of Cat�s Somatic Sensory Cortex”, Journal of Neurophysiology, 20:4,
pp. 408-434, 1957. - J. Szentagothai, “Architecture of the Cerebral Cortex”, in H. H. Jasper,
A. A. Ward and A Pope (eds.). Basic Mechanisms of the Epilepsies. Boston: Little, Brown, and Company, 1969. - J. Szentagothai, “Synaptology of the Visual Cortex”, in R. Jung (ed.). Handbook of Sensory Physiology, 7/3. New York: Springer-Verlag, 1973.
- A. E. Fessard, “Mechanisms of Nervous Integration and Conscious Experience”, in J. F. Delafreznaye (ed.). Brain Mechanisms and Consciousness. Springfield: C. C. Thomas, 1954.
- W. Penfield, “The Cerebral Cortex in Man, I, The Cerebral Cortex and Consciousness”, Archives of Neurology and Psychiatry, 40:3, pp. 417-442, 1938.
- W. Penfield, “Epileptic Automatism and the Centrencephalic Integrating System”, Research Publications: Association for Research in Nervous and Mental Disease, 30:4, pp. 513-528, 1952.
- W. Penfield, “Epilepsy, Neurophysiology and Some Brain Mechanisms Related to Consciousness”, in H. H. Jasper, A. A. Ward and A Pope (eds.). Basic Mechanisms of the Epilepsies. Boston: Little, Brown, and Company, 1969.
- W. Penfield and T. Rasmussen. The Cerebral Cortex in Man. New York: Macmillan, 1950.
- J. E. Bogen, “The Other Side of the Brain: An Appositional Mind”, in R. E. Ornstein (ed.). The Nature of Human Consciousness. New York: Viking Press, 1973.
- M. S. Gazzaniga, “The Split Brain in Man”, Scientific American, 217:2,
pp. 24-29, 1967. - M. S. Gazzaniga. The Bisected Brain. New York: Appleton-Century-Crofts, 1970.
- M. S. Gazzaniga, J. E. Bogen and R. W. Sperry, “Observations on Visual Perception After Disconnexion of the Cerebral Hemispheres in Man”, Brain, 88:Part 2, pp. 221-236, 1965.
- R. W. Sperry, “Cerebral Organization and Behavior”, Science, 133:3466,
pp. 1749-1757, 1961. - R. W. Sperry, “The Great Cerebral Commissure”, Scientific American, 210:1, pp. 42-45, 1964.
- R. W. Sperry, “Brain Bisection and Consciousness”, in J. C. Eccles (ed.). Brain and Conscious Experience. New York: Springer-Verlag, 1966.
- E. Jacobson. Biology of Emotions. Springfield: C. C. Thomas, 1967.
- E. Jacobson. Progressive Relaxation. Chicago: U. of Chicago Press, 1929.
- E. Jacobson, “Electrical Measurements of Neuromuscular States During Mental Activities: I. Imagination of Movement Involving Skeletal Muscle”, American Journal of Physiology, 91:2, pp. 567-608, 1930; “II. Imagination and Recollection of Various Muscular Acts”, ibid., 94:1, pp. 22-34, 1930;
“III. Visual Imagination and Recollection”, ibid., 95:3, pp. 694-702, 1930;
“IV. Evidence of Contraction of Specific Muscles During Imagination”, ibid., pp. 703-712; “V. Variation of Specific Muscles Contracting During Imagination”, ibid., 96:1, pp. 115-121, 1931; “VI. Note on Mental Activities Concerning an Amputated Limb”, ibid., pp. 122-125; “VII. Imagination, Recollection and Abstract Thinking, Involving the Speech Musculature”, ibid., 97:1, pp. 200-209, 1931. (Current comment by Derek Dillon: Though certainly not intended by Jacobson, this series of papers is the most profound body of scientific research ever produced on the mechanisms underlying efficacy of meditation and hatha yoga practice. Anyone engaging in a meditative discipline without awareness of the principles described here is surely receiving limited benefits from such practice.) - E. Jacobson, “Electrophysiology of Mental Activities”, American Journal of Psychology, 44, pp. 677-694, 1932.
- H. H. Jasper and C. Ajmone-Marsan, “Thalamocortical Integrating Mechanisms”, Research Publications: Association for Research in Nervous and Mental Disease, 30:4, pp. 493-512, 1952.
- H. H. Jasper, C. Ajmone and J. Stoll, “Cortico-fugal Projections to the Brain Stem”, Archives of Neurology and Psychiatry, 67:2, pp. 155-171, 1952.
- D. I. Mostofsky (ed.). Attention: Contemporary Theory and Analysis.
New York: Appleton-Century-Crofts, 1970. - S. Sharpless and H. H. Jasper, “Habituation of the Arousal Reaction”, Brain, 79: Part 4, pp. 655-680, 1956.
- G. Spencer Brown. Laws of Form. London: George Allen and Unwin, 1969.
- J. Hurwitz and J. T. August, “The Role of DNA in RNA Synthesis”, in
J. N. Davidson and W. E. Cohn (eds.). Progress in Nucleic Acid Research, 1. New York: Academic Press, 1963. - S. Bogoch. The Biochemistry of Memory. New York: Oxford U. Press, 1968.
- H. Hyden, “Biochemical Changes Accompanying Learning”, in G. C. Quarton, T. Melnechuk and F. O. Schmitt (eds.). The Neurosciences. New York: Rockefeller U. Press, 1967.
- H. Hyden, “Biochemical Aspects of Learning and Memory”, K. H. Pribram (ed.). On the Biology of Learning. New York: Harcourt, Brace and Company, 1969.
- H. Hyden, “Biochemical Changes in Glial Cells and Nerve Cells at Varying Activity”, in O. Hoffman-Ostenhof (ed.). Proceedings of the Fourth International Congress of Biochemistry, 3. New York: Pergamon Press, 1959.
- H. Hyden, “Introductory Remarks to the Session on Memory Processes”, Neurosciences Research Program Bulletin, 2:3, pp. 23-38, 1964.
- H. Hyden, “Some Brain Protein Changes Reflecting Neuronal Plasticity at Learning”, A. G. Karczmar and J. C. Eccles (eds.). Brain and Human Behavior. New York: Springer-Verlag, 1972.
- I. Tasaki and J. J. Chang, “Electric Response of Glia Cells in Cat Brain”, Science, 128:3333, pp. 1209-1210, 1958.
- Return to:
- •Top
- •The Ithaca Papers -- Abstracts
- •Home page
- •Top